
As you put the finishing touches on your paper, you notice the sun rising and fantasize about crawling in bed. Your vision and hearing are beginning to distort and the words staring back at you from the monitor have lost their meaning. Your brain … well, feels like mush. We’ve all been there. That debilitating brain fog that inevitably sets in after an all-nighter prompts the obvious question: what does sleep deprivation actually do to the brain?
Neuroscientists from Norway set out to answer this question in their recent PLOS ONE study, examining how a night forgoing sleep affects brain microstructure. Among their findings, sleep deprivation induced widespread structural alterations throughout the brain. The lead author shares his thoughts on the possible biological causes of these changes, and whether they may be long-lasting.
Inducing sleep deprivation
The researchers assessed a group of 21 healthy young men over the course of a day. The participants underwent diffusion tensor imaging (DTI; a form of MRI that measures water diffusion and can be used to evaluate white matter integrity) when they first awoke, at 7:30 am. They were free to go about their day as normal before returning for a second DTI scan at 9:30 pm. They remained in the lab for monitoring until a final scan at 6:30 am the following morning, for a total period of 23 hours of continued waking. Since we’re now learning that anything and everything can influence brain structure on surprisingly short time-scales, the researchers finely controlled as many confounding factors as possible. The participants were not allowed to exercise or consume alcohol, caffeine or nicotine during the study, or to eat right before the scans. Since DTI measures water diffusion, hydration was evaluated at all sessions and accounted for in their analysis.
Rapid microstructural changes to waking
The researchers were interested in two main questions: How does the brain change after a normal day of wakefulness and after sleep deprivation? They focused on three DTI metrics to probe how different features of neuronal tissue may change with waking. Radial diffusivity (RD) measures how water diffuses across fibres, whereas axial diffusivity (AD) measures diffusion along the length of a tract. Fractional anisotropy (FA) is the ratio of axial to radial diffusivity and therefore measures how strongly water diffuses along a single direction.
From morning to evening, FA increased and this was driven mostly by reduced RD (Figure, left). From the evening to the next morning — after the all-nighter — FA values decreased to levels comparable to the prior morning, and this drop was coupled with a decrease in AD (Figure, right). Thus, over the course of a full day of wakefulness FA fluctuated, temporarily rising but eventually rebounding. In contrast, both RD and AD declined but at different rates, RD dropping by the end of a normal day, and AD dropping later, only after considerable sleep deprivation. These changes were non-specific, occurring throughout the brain, including in the corpus callosum, brainstem, thalamus and frontotemporal and parieto-occipital tracts.
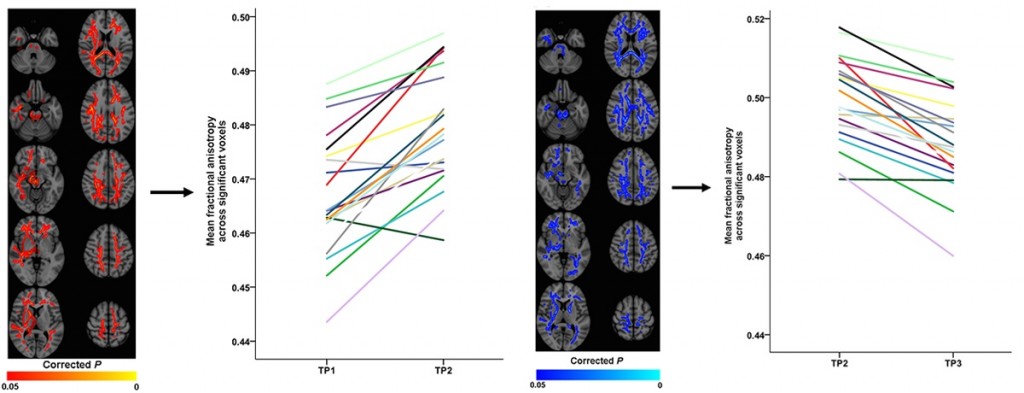
Throughout the brain, FA values increase from morning to evening (left) and decrease from the evening to the next morning after a night without sleep (right). Elvsåshagen et al., 2015.
How bad are the neuroconsequences of sleep deprivation?
Other studies have corroborated these reports that wakefulness alters the brain, including reduced diffusion with increasing time awake, and altered functional connectivity after sleep deprivation. How this plasticity reflects the consequences of waking on the brain, however, isn’t clear. Sleep is known to be essential to tissue repair and is particularly important for promoting lipid integrity to maintain healthy cell membranes and myelination. The question remains, therefore, how detrimental the structural reorganization from sleep deprivation really is. Does the plasticity reported here and elsewhere persist for days, weeks or longer, or can a long night of deep catch-up sleep reverse any detriment that all-nighter caused?
“My hypothesis,” says first author Dr. Torbjørn Elvsåshagen, “would be that the putative effects of one night of sleep deprivation on white matter microstructure are short term and reverse after one to a few nights of normal sleep. However, it could be hypothesized that chronic sleep insufficiency might lead to longer-lasting alterations in brain structure. Consistent with this idea, evidence for an association between impaired sleep and localised cortical thinning was found in obstructive sleep apnea syndrome, idiopathic rapid eye movement sleep behaviour disorder, mild cognitive impairment and community-dwelling adults. Whether chronic sleep insufficiency can lead to longer-lasting alterations in white matter structure remains to be clarified.”
Is sleepiness really to blame?
It’s likely that multiple factors contribute to these distinct patterns of change in neuronal tissue. After sleep deprivation, the extent of AD decline correlated with subjective sleepiness ratings, suggesting that microstructural alterations may in fact be attributable to changes in alertness or arousal. This possibility is in line with the finding that changes occurred in both the thalamus and brainstem, regions important for arousal and wakefulness. However, the non-linear changes in FA suggest that some microstructural changes may be less related to sleepiness and more directly driven by circadian effects. FA increased late in the day, but — despite fatigue — dropped back after sleep deprivation to the same levels as the day prior. This rebounding may have been due to declining levels of AD and RD reaching equilibrium (reminder, FA is the ratio of AD to RD) or to neuronal features that fluctuate with our circadian rhythms, at least partially independent of our sleep habits. What’s more, other studies have found that presumably mundane activities, for example juggling or spatial learning, also induce grey and white matter changes within hours, and presumably many more as-of-yet unstudied activities also cause similarly rapid plasticity. Given that participants were free to engage in various physical and cognitive activities between the scans, it’s reasonable to assume that some of these behaviours may have also influenced brain structure. Whatever the mechanism, these effects underscore the importance of accounting for time of day in structural neuroimaging studies.
Dr. Elvsåshagen elaborates on these possible factors: “The precise neurobiological substrate for the observed DTI changes after waking remain to be clarified. We cannot rule out the possibility that both activity-independent and activity-dependent processes could contribute to DTI changes after waking. In support of potential activity-dependent white matter alterations, there is interesting evidence from in vitro studies indicating that hours of electrical activity can lead to changes in myelination. To further explore the possibility of activity-dependent white matter alterations, one could examine whether different physical or cognitive tasks lead to task-specific white matter changes.”
Sleepy outliers?
Notably, two of the 21 participants did not show the same rise in FA throughout the day as the others, and showed the smallest change in FA and AD after sleep deprivation. While variability across individuals in terms of brain structure and biological responses to the environment is expected, the remarkable consistency of the study’s other findings raises the possibility that some other factors may explain these outliers. Dr. Elvsåshagen conjectures, “These individuals were also the least tired individuals after sleep deprivation. Although highly speculative, one possible explanation for the lesser changes in these two participants might be a particular resistance to the effects of waking and sleep deprivation.” A follow-up with additional time-points and closer monitoring of activities could help more finely track how the patterns of brain microstructural change shift over periods of waking, and vary across individuals.
Linking diffusion to neurons
How sleep, fatigue, activity or circadian rhythms affect particular neuronal structural properties remains to be seen. RD and AD are thought to depend onmyelin and axon integrity, respectively, but DTI metrics in general are sensitive to various other tissue features as well, including cell membrane permeability, axon diameter, tissue perfusion or glial processes. While these properties are difficult to image in living humans, insight from animal studies will help determine how waking impacts specific neuronal characteristics.
Longer-term studies are needed to answer these questions. Dr. Elvsåshagen shared that his team has since replicated their results in a larger sample, and are planning a follow-up study on the effects of waking and sleep deprivation on grey matter structure. Until these outstanding questions are answered, keeping a regular sleep schedule — and avoiding those early morning paper-writing marathons — may be better option for your brain health.
Any views expressed are those of the author, and do not necessarily reflect those of PLOS.
Emilie Reas received her PhD in Neuroscience from UC San Diego, where she used fMRI to study memory. As a postdoc at UCSD, she currently studies how the brain changes with ageing and disease. In addition to her tweets for @PLOSNeuro she is @etreas.
Top image by Vladimir Agafonkin under Creative Commons licence.
References
Bellesi M, Pfister-Genskow M, Maret S, Keles S, Tononi G, Cirelli C (2013). Effects of sleep and wake on oligodendrocytes and their precursors. J Neurosci. 33: 14288 — 14300. doi: 10.1523/JNEUROSCI.5102-12.2013
Budde MD, Xie M, Cross AH, Song SK (2009). Axial diffusivity is the primary correlate of axonal injury in the experimental autoimmune encephalomyelitis spinal cord: a quantitative pixelwise analysis. J Neurosci. 29: 2805 — 2813. doi: 10.1523/JNEUROSCI.4605-08.2009
De Havas JA, Parimal S, Soon CS, Chee MW (2012). Sleep deprivation reduces default mode network connectivity and anti-correlation during rest and task performance. NeuroImage. 59: 1745 — 1751. doi: 10.1016/j.neuroimage.2011.08.026
Driemeyer J, Boyke J, Gaser C, Buchel C, May A (2008). Changes in grey matter induced by learning — revisited. PLOS ONE. 3: e2669. doi: 10.1371/journal.pone.0002669
Elvsåshagen T, Norbom LB, Pedersen PØ, Quraishi SH, Bjørnerud A, Malt UK (2015). Widespread Changes in White Matter Microstructure after a Day of Waking and Sleep Deprivation. PLOS ONE. 10(5): e0124859. doi: 10.1371/journal.pone.0127351
Hinard V, et al. (2012). Key electrophysiological, molecular, and metabolic signatures of sleep and wakefulness revealed in primary cortical cultures. J Neurosci. 32: 12506 — 12517. doi: 10.1523/JNEUROSCI.2306-12.2012
Hofstetter S, Tavor I, Tzur Moryosef S, Assaf Y (2013). Short-term learning induces white matter plasticity in the fornix. J Neurosci. 33: 12844 — 12850. doi: 10.1523/JNEUROSCI.4520-12.2013
Jiang C, , et al. (2014). Diurnal microstructural variations in healthy adult brain revealed by diffusion tensor imaging. PLOS ONE. 9: e84822. doi: 10.1371/journal.pone.0084822
Joo EY, et al. (2013). Brain Grey Matter Deficits in Patients with Chronic Primary Insomnia. Sleep. 36(7): 999-1007. doi: 10.5665/sleep.2796
Rayayel S, et al. (2015). Patterns of cortical thinning in idiopathic rapid eye movement sleep behaviour disorder. Mov Disord. 30(5): 680 — 687. doi: 10.1002/mds.25820
Sanchez-Espinosa MP, Atienza M, Cantero JL (2014). Sleep deficits in mild cognitive impairment are related to increased levels of plasma amyloid-β and cortical thinning. NeuroImage. 98: 395-404. doi: 10.1016/j.neuroimage.2014.05.027
Song SK, Sun SW, Ramsbottom MJ, Chang C, Russell J, Cross AH (2002). Dysmyelination revealed through MRI as increased radial (but unchanged axial) diffusion of water. NeuroImage. 17: 1429 — 1436. doi: 10.1006/nimg.2002.1267
Sexton CE, et al. (2014). Accelerated changes in white matter microstructure during ageing: a longitudinal diffusion tensor imaging study. J Neurosci. 34(46): 15425 — 15436. doi: 10.1523/JNEUROSCI.0203-14.2014
Wake H, Lee PR, Fields RD (2011). Control of Local Protein Synthesis and Initial Events in Myelination by Action Potentials. Science. 333(6049): 1647 — 1651. doi: 10.1126/science.1206998
.